






TRANSLACJA (zobacz i por≤wnaj: translacja u Procaryota)
Translacja - proces, w kt≤rym nastΩpuje odczyt informacji genetycznejz mRNA i synteza bia│ka. Bior▒ w nim udzia│ opr≤cz matrycy ( mRNA ) i aminokwas≤w tak┐e cz▒steczki tRNA ( dostarczaj▒ce aminokwas≤w ), rybosomy oraz szereg czynnik≤w wspomagaj▒cych.
Przetransportowany do cytoplazmy mRNA mo┐e ulec translacji, b▒d╝ te┐ zostaµ szybko zdegradowany, je╢li bia│ko jakie jest przeze± zakodowane wystΩpuje w kom≤rce w dostatecznej ilo╢ci. I tak na przyk│ad mRNA dla histon≤w jest bardzo stabilne w fazie S cyklu kom≤rkowego, tj.
w tym momencie, kiedy obserwuje siΩ najwy┐sze zapotrzebowanie na histony. Zatem w fazie S nastΩpuje translacja mRNA histonowego. W innych fazach cyklu kom≤rkowego stabilno╢µ tych transkrypt≤w jest ok. 5-krotnie ni┐sza, co oznacza, ┐e ulegaj▒ one degradacji.
Wniosek z tego, ┐e synteza bia│ka zachodzi w zale┐no╢ci od potrzeb kom≤rki, a zatem regulacja ekspresji informacji genetycznej odbywa siΩ tak┐e na etapie poprzedzaj▒cym translacjΩ.
Rybosomy
(zobacz i por≤wnaj: rybosomy prokariotyczne)Miejscem syntezy bia│ka s▒ organella kom≤rkowe zwane rybosomami. WystΩpuj▒ one opr≤cz cytoplazmy tak┐e w mitochondriach i chloroplastach, ale te rodzaje rybosom≤w nie bΩd▒ tematem naszych rozwa┐a±. Rybosomy w cytoplazmie mog▒ wystΩpowaµ w formie wolnej lub zwi▒zanej z retikulum endoplazmatycznym. Pierwsze z nich uczestnicz▒ w translacji bia│ek cytosolowych, a drugie bia│ek eksportowanych do wnΩtrza retikulum oraz na zewn▒trz kom≤rki. Obydwa rodzaje rybosom≤w zbudowane s▒ z dw≤ch podjednostek:
- ma│ej, kt≤rej sta│a sedymentacji wynosi 40S;
- du┐ej o wsp≤│czynniku sedymentacji r≤wnym 60S.
Sk│adaj▒ siΩ one z rybosomalnego RNA ( 18S rRNA ma│ej podjednostki; 5, 5.8, 26 ( 28S rRNA du┐ej podjednostki ) oraz kilkudziesiΩciu bia│ek, kt≤rych sk│ad r≤┐ni siΩ miΩdzy obydwiema "czΩ╢ciami" rybosomu. W obrΩbie rybosomu wyr≤┐nia siΩ trzy istotne miejsca zwane E ( exit - tΩdy wychodzi "zu┐yty" tRNA ), A ( aminokwasowe ), P ( peptydowe ), o kt≤rych bΩdzie mowa w dalszych czΩ╢ciach tego rozdzia│u.
Przebieg translacji
Translacja mRNA zachodzi w kierunku 5'« 3', zatem pierwszy aminokwas syntetyzowanego polipeptydu zakodowany jest przez kodon ( patrz Kod genetyczny ) le┐▒cy bli┐ej 5' ko±ca transkryptu. Natomiast najwcze╢niej wbudowanymi aminokwasami powstaj▒cego bia│ka s▒ te, kt≤re stanowi▒ jego N - terminaln▒ czΩ╢µ. Wynika to z mechanizmu tworzenia wi▒za± peptydowych miΩdzy sasiadujacymi aminokwasami ( patrz "Chemiczne podstawy biologii molekularnej" ).
TranslacjΩ podzieliµ mo┐na na trzy zasadnicze etapy:
- inicjacjΩ,
- elongacjΩ,
- terminacjΩ.
Inicjacja
Pierwszym etapem syntezy bia│ka jest przy│▒czenie mRNA do mniejszej podjednostki rybosomu ( 40S ) w obecno╢ci inicjatorowego tRNA i szeregu czynnik≤w inicjuj▒cych.
U eukariont≤w pierwszym kodonem ulegaj▒cym translacji jest kodon AUG ( tzw. kodon start ), koduj▒cy metioninΩ.
Poniewa┐ w mRNA tr≤jek takich mo┐e byµ bardzo wiele, miejsce pocz▒tku translacji wyznacza ten kodon, kt≤ry po│o┐ony jest najbli┐ej 5' ko±ca. W poszukiwaniu pierwszego tj. inicjacyjnego kodonu metioninowego bior▒ udzia│ czynniki zwane eIF3 i eIF4, z kt≤rych pierwszy przy│▒cza do struktury czapeczki kompleks bia│kowy ( tzw. CBP ) oraz odszukuje AUG, natomiast drugi - eIF4 dostarcza energii do poszukiwa±. Zanim jednak rybosom ze zwi▒zanym bia│kiem eIF3 zacznie kroczyµ po mRNA w poszukiwaniu AUG, do podjednostki 40S zwiazany zostaje inicjatorowy aminoacylo tRNA tj. tRNA nios▒cy cz▒steczkΩ aminokwasu - tu metioniny. Gdy przesuwaj▒ca siΩ po mRNA podjednostka 40S ze zwi▒zanym Met-tRNA natrafi na kodon "start" nastΩpuje przy│▒czenie wiΩkszej podjednostki rybosomu ( 60S ).
|
Nazwa czynnika |
Funkcje |
|
eIF2 |
Przy│▒czenie do podjednostki 40S inicjacyjnego aminoacylo tRNA |
|
eIF3 |
Udzia│ w przy│▒czaniu do 5Æ ko±ca mRNA bia│ek CBP Odszukiwanie kodonu inicjacyjnego Uniemo┐liwianie asocjacji podjednostek rybosomowych pod nieobecno╢µ innych czynnik≤w inicjacyjnych |
|
eIF4 |
Dostarczanie energii niezbΩdnej w dzia│aniu eIF3 z hydrolizy ATP |
|
eIF5 |
Indukcja uwolnienia eIF2 i eIF3 kosztem energii z hydrolizy GTP ( zwi▒zanego z eIF2 ) |
Przk│ady eukariotycznych czynnik≤w inicjacyjnych
Inicjacja translacji stanowi kolejny punkt kontrolny w ekspresji informacji genetycznej. Regulacja ekspresji na tym etapie odbywa siΩ za po╢rednictwem czynnika eIF2, kt≤ry zdolny jest do inicjowania translacji tylko wtedy, gdy posiada przy│▒czon▒ cz▒steczkΩ GTP. W nieaktywnym eIF2 przy│▒czony jest GDP, kt≤ry mo┐e byµ wymieniony na GTP za po╢rednictwem bia│ka GEF, w wyniku reakcji:
Reakcja ta jednak mo┐e byµ zaburzona wskutek fosforylacji eIF2 nieaktywnego, w kt≤rym niemo┐liwa jest wymiana nukleotydu guanylowego ( GDP/GTP ). Innymi s│owy, fosforylacja skazuje bia│ko eIF2 na brak aktywno╢ci.
Elongacja │a±cucha polipeptydowego
Po z│o┐eniu rybosomu, tj. przy│▒czeniu podjednostki 60S do powsta│ego uk│adu, przez odpowiednie tRNA dostarczane s▒ kolejne aminokwasy. Rybosom przesuwa siΩ stopniowo wzd│u┐ nici mRNA, do kt≤rej na zasadzie komplementarno╢ci dopasowuj▒ siΩ cz▒steczki aminoacylo tRNA. Dopasowanie to dotyczy obszaru antykodonu tj. tr≤jki nukleotyd≤w (z tRNA) "pasuj▒cych" do kodonu z mRNA. I tak z kodonem CAC oddzia│ywaµ bΩdzie tRNA zawieraj▒cy antykodon GUG i nios▒cy histydynΩ.
Do kolejnego aminokwasu dostarczonego w miejsce A rybosomu przez aminoacylo tRNA przy│▒cza siΩ za po╢rednictwem grupy karboksylowej wcze╢niej odczytany aminokwas wystΩpuj▒cy w danym momencie - jeszcze w postaci zwi▒zanej z tRNA - w miejscu P rybosomu. Tzn. je╢li pierwszym kodonem jest AUG, drugim CAC, a trzecim UUU bΩdziemy mieli do czynienia z nastΩpuj▒cymi zmianami na terenie rybosomu:
1. Do dostarczonej jako drugiej w kolejno╢ci histydyny przy│▒cza siΩ inicjacyjna metionina. Na tRNA histydynowym ( w miejscu A ) tworzy siΩ dipeptyd Met-His i przesuwa siΩ w miejsce P.
2. Do przy│▒czonej jako trzeciej fenyloalaniny grup▒ karboksylow▒ przy│▒czy siΩ dipeptyd Met-His i powstanie tr≤jpeptyd Met-His-Phe, przemieszczany jak poprzednio w miejsce P.
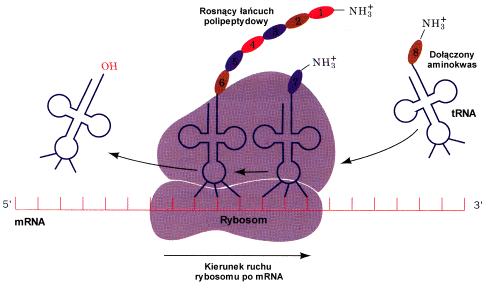
Og≤lny schemat translacji
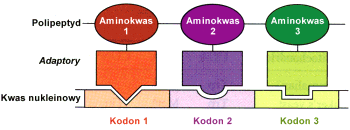
Teoria adaptorowa dotycz▒ca rozpoznawania kodon≤w
Tak wiΩc, wyd│u┐anie siΩ │a±cucha polipeptydowego polega na przy│▒czaniu kolejnych aminokwas≤w na jego C-ko±cu. ElongacjΩ │a±cucha polipeptydowego umo┐liwiaj▒ tzw. czynniki elongacyjne m.in. eEF1( i eEF1( odpowiedzialne za dostarczanie aminoacylo tRNA, oraz eEF2 odpowiedzalny za translokacjΩ tj. przeniesienie z m-ca A na m-ce P odbywaj▒c▒ siΩ kosztem energii z GTP.
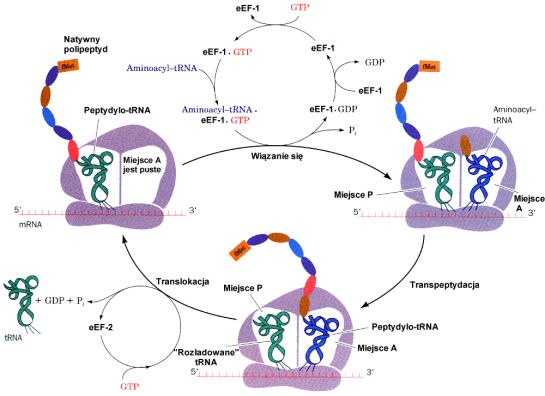
Cykl elongacyjny
Terminacja
Translacja ko±czona jest w miejscu, w kt≤rym wyst▒pi jeden spo╢r≤d trzech kodon≤w terminacyjnych tzw. kodon≤w "stop" ( UAG, UGA, UAA ). Wymienione tr≤jki nukleotydowe rozpoznawane s▒ w odr≤┐nieniu do ca│ej reszty nie przez amino-acylo tRNA, a przez tzw. czynnik uwalniaj▒cy eRF. Czynnik ten aktywuje enzym rozcinaj▒cy wi▒zanie miΩdzy polipeptydem a cz▒steczk▒ tRNA, kt≤ra dostarczy│a ostatniego aminokwasu, czego skutkiem jest uwolnienie │a±cucha polipeptydowego. Zdarzenie to jest uwie±czeniem szeregu proces≤w, jakie obejmuje ekspresjia informacji genetycznej.
Podsumowuj▒c:
1. Translacja zachodzi w rybosomach - strukturach rybonukleoproteinowych (RNA : bia│ko );
2. Sygna│em pocz▒tku syntezy bia│ka jest kodon AUG, natomiast ko±ca syntezy jeden z kodon≤w stop - UAA, UAG, UGA;
3. Synteza bia│ka wspomagana jest przez czynniki bia│kowe tzw. : inicjacyjne, elongacyjne i terminacyjne;
4. Translacja jest procesem energoch│onnym, czerpi▒cym energiΩ z hydrolizy GTP i ATP;
 OBR╙BKA PRE-mRNA
OBR╙BKA PRE-mRNA

⌐ 1997, 1998 Biologia Molekularna w Internecie Webmaster