






SOMATYCZNE REARAN»ACJE W UKúADZIE IMMUNOLOGICZNYM
Unikalne zjawiska tego typu zachodz▒ w uk│adzie immunologicznym (odporno╢ciowym) krΩgowc≤w. Jego podstawow▒ funkcj▒ jest obrona organizmu przed atakuj▒cymi go mikroorganizmami. Musi szybko i sprawnie rozpoznaµ zagra┐aj▒cy organizmowi patogen i zlik widowaµ go.
Mo┐na sobie wyobraziµ, jak niezwykle trudne jest to zadanie.
Znamy ogromnie wiele patogen≤w i wci▒┐ odkrywa siΩ nowe. Wirusy, grzyby, pierwotniaki, robaki ...
Na dodatek mikroorganizmy ewoluuj▒ najszybciej spo╢r≤d ca│ego ╢wiata ┐ywego (link - ewolucja molekularna) Niezwykle wysokie tempo tych przemian podnosi jeszcze wymagania stawiane uk│adowi immunologicznemu, zmuszaj▒c go do swoistej "pogoni" za mutuj ▒cymi patogenami. Musi on byµ przygotowany nawet na odparcie ataku nie istniej▒cych jeszcze szczep≤w bakterii czy wirus≤w.
Biolog≤w od dawna intrygowa│a ta zdolno╢µ organizmu do odpowiedzi na praktycznie nieograniczon▒ liczbΩ antygen≤w.
Sekret skutecznej obrony przed inwazj▒ patogen≤w tkwi przede wszystkim w szybkiej i precyzyjnej identyfikacji przeciwnika. S│u┐▒ temu miΩdzy innymi przeciwcia│a, czyli immunoglobuliny (Ig). Ka┐de przeciwcia│o posiada zdolno╢µ do wi▒zania specyficzn ego dla siebie antygenu.
Skoro jednak organizmowi zagra┐a tak wiele r≤┐nych mikroorganizm≤w, to jak nasz uk│ad immunologiczny jest w stanie wytworzyµ odpowiedni▒ ilo╢µ przeciwcia│ ?
Pr≤bowa│a to wyja╢niµ "teoria informacyjna" Paulinga i Horowitza, kt≤ra zak│ada│a, ┐e organizm, kt≤ry nie zetkn▒│ siΩ jeszcze z danym antygenem, nie zawiera kom≤rek ani przeciwcia│ zdolnych do jego swoistego rozpoznania. Zgodnie z t▒ hipotez▒ antyg en odkszta│ca│by nieczynn▒ immunoglobulinΩ, aktywuj▒c j▒ i dostosowuj▒c do swojej struktury przestrzennej. Rozwa┐ano te┐ mo┐liwo╢µ oddzia│ywania antygenu na gen koduj▒cy │a±cuchy bia│kowe przeciwcia│a.
Teoria ta okaza│a siΩ jednak sprzeczna z rezultatami do╢wiadcze±. Wykazano w nich, miΩdzy innymi, ┐e w mieszaninie limfocyt≤w B pobranych od dawcy, kt≤ry nigdy nie zetkn▒│ siΩ z danym antygenem, znajduj▒ siΩ kom≤rki zdolne do swoistego rozpoznania tego w│a╢nie antygenu i produkcji odpowiednich przeciwcia│ !
P≤╝niej zbadano, ┐e organizm wytwarza oko│o 108 - 109
(1 000 000 000 ) rodzaj≤w przeciwcia│ (!). Liczba ta jest jeszcze bardziej zaskakuj▒ca, gdy u╢wiadomimy sobie, ┐e ca│y genom cz│owieka zawiera prawdopodobnie tylko oko│o 105 (100 000) gen≤w koduj▒cych bia│ka! Prosty schemat syntezy bia│ka: DNA « RN A « BIAúKO nie dostarcza odpowiedzi na pytanie o ╝r≤d│o takiej r≤┐norodno╢ci immunoglobulin. Wyt│umaczeniem okaza│o siΩ odkrycie somatycznych rearan┐acji genomu limfocyt≤w odpowiedzialnych za wytwarzanie immunoglobulin.
Aby zrozumieµ ten niezwyk│y proces przyjrzyjmy siΩ najpierw budowie pojedynczego przeciwcia│a.
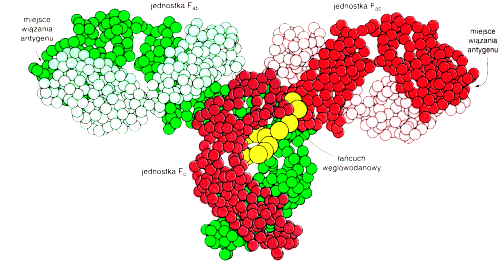
Dla u│atwienia immunoglobulinΩ przedstawiamy schematycznie w kszta│cie litery "Y".
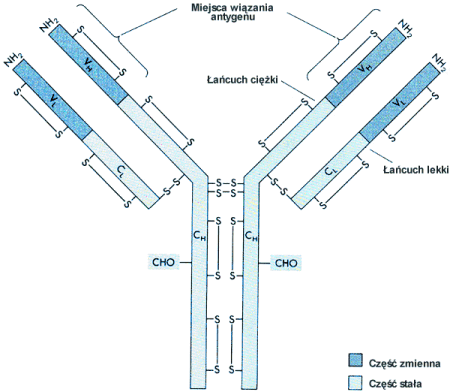
Fragment Fc przeciwcia│a umo┐liwia │▒czenie siΩ immunoglobulin w wiΩksze agregaty - dimery, pentamery itp., a tak┐e kotwiczenie przeciwcia│a w b│onie kom≤rkowej. Jest te┐ odpowiedzialny za aktywowanie innych sk│adnik≤w odpowiedzi immunologicznej. F ragment Fab (antigen binding) immunoglobuliny odpowiada za specyficzne wi▒zanie antygenu.
Geny koduj▒ce │a±cuchy polipeptydowe sk│adaj▒ce siΩ na przeciwcia│o znajduj▒ siΩ u cz│owieka na chromosomach 2, 14 i 22.
Procesy, kt≤re prowadz▒ do tak niezwyk│ej r≤┐norodno╢ci przeciwcia│, najlepiej mo┐na przedstawiµ na przyk│adzie syntezy │a±cucha ciΩ┐kiego Ig, kt≤rego gen u cz│owieka umiejscowiony jest na chromosomie 14.
Ogromna wiΩkszo╢µ gen≤w Eucaryota jest nieci▒g│a (linki do rozdz. Eucaryota). Tak┐e i geny koduj▒ce immunoglobuliny nie s▒ tu wyj▒tkiem. Egzony koduj▒ce czΩ╢ci │a±cucha polipeptydowego porozdzielane s▒ fragmentami DNA nie nios▒cego ┐adnej infor macji (a osi▒gaj▒cymi d│ugo╢µ do 8 tysiΩcy par zasad !). Jednak to nie nieci▒g│o╢µ genu stanowi o unikalnym charakterze zjawiska syntezy przeciwcia│.
Niezwyk│▒ cech▒ genu │a±cuch≤w przeciwcia│ jest wystΩpowanie obok siebie wielu, nieznacznie siΩ od siebie r≤┐ni▒cych, kopii danego egzonu.
Gen fragmentu ciΩ┐kiego immunoglobuliny wygl▒daµ bΩdzie mniej wiΩcej tak:
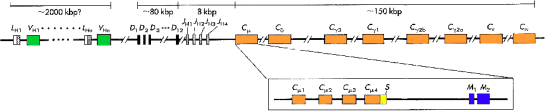
Forma genu dla │a±cucha ciΩ┐kiego immunoglobuliny, pokazana powy┐ej, nie wystΩpuje jednak w dojrza│ych limfocytach, zdolnych do wytwarzania Ig. Znale╝µ j▒ mo┐na miΩdzy innymi w kom≤rkach p│ciowych i niezr≤┐nicowanych kom≤rkach macierzystych, st▒d t a forma genu nosi nazwΩ zarodkowej.
W dojrzewaj▒cym limfocycie B zachodzi niezwyk│y proces somatycznej rekombinacji polegaj▒cy na przegrupowaniach w DNA koduj▒cym │a±cuch Ig. Rekombinacje te prowadz▒ do wytworzenia funkcjonalnych gen≤w immunoglobulinowych.
Pierwsze przegrupowanie w obrΩbie obszaru koduj▒cego │a±cuch ciΩ┐ki przeciwcia│a prowadzi do po│▒czenia jednego z segment≤w D z jednym z segment≤w J. Odbywa siΩ to najczΩ╢ciej poprzez wypΩtlenie nici i delecjΩ fragmentu DNA dziel▒cego │▒czone segme nty.
W nastΩpnym etapie dochodzi do │▒czenia powsta│ej sekwencji DJ z jednym z egzon≤w z grupy V.
Wycinanie odcink≤w DNA pomiΩdzy segmentami i │▒czenie odpowiednich segment≤w zachodzi przy udziale rekombinaz - s│abo dot▒d poznanych enzym≤w, aktywnych jedynie w rozwijaj▒cych siΩ limfocytach.
Rekombinaza rozpoznaje tzw. "sekwencje sygna│owe" znajduj▒ce siΩ na ko±cach segment≤w (egzon≤w). Umo┐liwia to precyzyjne po│▒czenie siΩ tylko jednego segmentu D z jednym z segment≤w J, jednym z segment≤w V, a p≤╝niej z jednym z segment≤w grupy C, d ecyduj▒cym o przynale┐no╢ci do klasy przeciwcia│.
Dodatkow▒ ciekawostk▒ mo┐e byµ fakt, ┐e egzony z poszczeg≤lnych rodzin (V,D,J), nie s▒ wybierane ca│kowicie losowo, tak, jak to pocz▒tkowo przypuszczano. W procesie rekombinacji preferowane s▒ np. egzony V le┐▒ce bli┐ej segment≤w J (u cz│owieka prz yk│adem mog▒ byµ segmenty VH6
i VH26) Podobne zjawisko zaobserwowano dla segment≤w J i D. Przyczyna takiego zjawiska, ograniczaj▒cego przecie┐ r≤┐norodno╢µ produkowanych przeciwcia│, nie zosta│a dot▒d dok│adnie zbadana. Przypuszcza siΩ jednak, ┐e preferowane kombinacje egzon≤w umo┐ liwiaj▒ wytworzenie przeciwcia│ o kluczowej dla bezpiecze±stwa organizmu swoisto╢ci.
Om≤wione do tej pory mechanizmy prowadz▒ce do r≤┐norodno╢ci przeciwcia│ pozwalaj▒ na wytworzenie najwy┐ej oko│o 105 Ig o r≤┐nej swoisto╢ci. Znacznie wiΩksze rzeczywiste bogactwo immunoglobulin ma swe ╝r≤d│o w innych jeszcze procesach. Jednym z bard ziej niezwyk│ych jest zmienno╢µ na z│▒czach.
Ten typ zmienno╢ci jest rezultatem braku precyzji przy │▒czeniu poszczeg≤lnych segment≤w V, D i J i przy wycinaniu znajduj▒cego siΩ pomiΩdzy nimi DNA. Przy formowaniu z│▒cza pomiΩdzy egzonami mo┐e doj╢µ do delecji nawet do 20 nukleotyd≤w. Ale nie t o jest najciekawsze.
Na z│▒czach miΩdzy sk│adanymi segmentami dochodzi bowiem do insercji dodatkowych (do 15) nukleotyd≤w. Proces do│▒czania nowych nukleotyd≤w ZACHODZI BEZMATRYCOWO !!! Oznacza to tworzenie "nowego genu", nie istniej▒cego w kom≤rce zarodkowej, w spos≤b ca│kowicie przypadkowy - "z powietrza" ! Za to niesamowite zjawisko odpowiedzialna jest transferaza nukleotyd≤w terminalnych (TdT). Tak powsta│e odcinki DNA na z│▒czach nazywamy regionami N.
Unikalnym procesem, zwiΩkszaj▒cym jeszcze liczbΩ wariant≤w Ig, jest przy│▒czanie do z│▒cza kr≤tkich (5-7 bp) oligonukleotyd≤w wycinanych podczas innych rekombinacji. To zjawisko bywa nazywane "chwytaniem oligonukleotyd≤w".
Szacuje siΩ, ┐e procesy zwi▒zane ze zmienno╢ci▒ na z│▒czach zwiΩkszaj▒ liczbΩ wariant≤w Ig o dalsze dwa rzΩdy wielko╢ci.
Nasz uk│ad immunologiczny, w obronie przed patogenami, wytworzy│ jeszcze inne metody zwiΩkszaj▒ce pulΩ produkowanych przeciwcia│.
Bardzo wydajnym procesem s▒ mutacje somatyczne. Zachodz▒ one we fragmentach genu koduj▒cych rejon zmienny │a±cucha ciΩ┐kiego i lekkiego Ig. Do indukcji mutacji dochodzi w limfocycie B pobudzonym przez kontakt z antygenem. Omawiane mutacje s▒ najczΩ ╢ciej mutacjami punktowymi, rzadziej zdarzaj▒ siΩ delecje czy insercje. Jedn▒ mutacjΩ obserwuje siΩ na ok. 10 000 kom≤rek spo╢r≤d jednej generacji. Jest to czΩstotliwo╢µ olbrzymia, oko│o milion razy wiΩksza ni┐ w przypadku mutacji spontanicznych w innych genach ! W wyniku tego procesu dochodzi do wymiany nawet do 1% nukleotyd≤w w egzonach dla czΩ╢ci zmiennych ciΩ┐kich i lekkich │a±cuch≤w Ig.
Mutacje te mog▒ prowadziµ do zwiΩkszenia powinowactwa do antygenu nawet o 100 razy i zwiΩkszaj▒ liczbΩ wariant≤w o kilka rzΩd≤w wielko╢ci !
Na ogromn▒ r≤┐norodno╢µ wariant≤w przeciwcia│ wp│ywa r≤wnie┐ fakt, ┐e poza │a±cuchami ciΩ┐kimi, kt≤rych syntezΩ opisali╢my powy┐ej, sk│adaj▒ siΩ one r≤wnie┐ z │a±cuch≤w lekkich, przy kt≤rych powstawaniu zachodz▒ niezale┐nie podobne procesy.
DziΩki tak wyrafinowanym metodom ekspresji gen≤w Ig, organizm mo┐e wytwarzaµ imponuj▒c▒ liczbΩ przeciwcia│ o r≤┐nej swoisto╢ci. Przy wykorzystaniu stosunkowo skromnej liczby gen≤w immunoglobulinowych, pula przeciwcia│ mog│aby osi▒gn▒µ oko│o 1011 =1 00 000 000 000 ! Liczba ta mog│aby jeszcze wzrosn▒µ, gdy┐ przy rachunku nie wziΩto pod uwagΩ trudnej do oszacowania, a niezwykle wydajnej, zmienno╢ci mutacyjnej.
Uk│ad immunologiczny cz│owieka wypracowa│ sobie wydajny system obrony przed zagro┐eniem ze strony chorobotw≤rczych mikroorganizm≤w. P│aci jednak wysok▒ cenΩ za zwyciΩstwo w wy╢cigu z niezwykle zr≤┐nicowanym i szybko mutuj▒cym przeciwnikiem. Niezwyk le drastyczne i ryzykowne metody: indukowanie mutacji, niedok│adny splicing, czy bezmatrycowa synteza DNA musz▒ wywo│ywaµ r≤wnie┐ negatywne skutki.
Szacuje siΩ, ┐e oko│o 80% rekombinacji powsta│ych dziΩki zmienno╢ci na z│▒czach jest nieproduktywne (!) tj. nie prowadzi do syntezy czynnego │a±cucha. Zmienia siΩ bowiem ramka odczytu, triplety nukleotyd≤w koduj▒ inne aminokwasy, lub powoduj▒ powstawanie kr≤tszego produktu. Somatyczne mutacje mog▒ r≤wnie dobrze zwiΩkszaµ powinowactwo (si│Ω wi▒zania antygenu), jak i prowadziµ do ca│kowitej jego utraty.
Powstanie limfocytu produkuj▒cego nieprawid│owe, nieczynne przeciwcia│a, lub nie wytwarzaj▒cego ich wcale nie jest jeszcze tak niebezpieczne. Mo┐e bowiem doj╢µ do sytuacji, ┐e limfocyt, na skutek somatycznych rearan┐acji zacznie wytwarzaµ przeciwci a│a skierowane przeciwko w│asnym antygenom!
W normalnych warunkach, jeszcze w ┐yciu p│odowym, zachodzi selekcja i eliminacja limfocyt≤w B produkuj▒cych przeciwcia│a rozpoznaj▒ce w│asne antygeny kom≤rkowe. Proces ten odbywa siΩ w szpiku kostnym. Podobne zjawiska, zachodz▒ce w grasicy, powoduj ▒ likwidacjΩ limfocyt≤w T zdolnych do atakowania antygen≤w w│asnego organizmu.
Jednak na skutek niedok│adnej eliminacji wymienionych powy┐ej kom≤rek, lub podczas pobudzenia uk│adu immunologicznego wywo│anego infekcj▒, uk│ad odporno╢ciowy mo┐e zwr≤ciµ siΩ przeciwko w│asnym kom≤rkom i tkankom.
Proces ten, nazywany autoagresj▒, mo┐e prowadziµ do ciΩ┐kich chor≤b autoimmunologicznych. Przyk│adem takich schorze± mo┐e byµ choroba Gravesa-Basedova, w kt≤rej autoantygenem s▒ receptory dla tyreotropiny zlokalizowane w tarczycy. W stwardnieniu ro zsianym organizm produkuje autoprzeciwcia│a skierowane przeciwko zasadowemu bia│ku mieliny, tworz▒cemu os│onki kom≤rek nerwowych. CzΩst▒ chorob▒ autoimmunologiczn▒ jest cukrzyca typu I, w kt≤rej organizm atakuje w│asne kom≤rki ( trzustki.
R≤wnie┐ niekt≤re nowotwory s▒ cen▒ p│acon▒ za sprawno╢µ uk│adu odporno╢ciowego przyk│adem mo┐e byµ ch│oniak Burkitta, choroba wystΩpuj▒ca najczΩ╢ciej w Afryce, w kt≤rej dochodzi do przeniesienia protoonkogenu c -myc (tu 2 oddzielne linki) z 8 chrom osomu, w niezwykle aktywne transkrypcyjnie miejsce gen≤w immunoglobulinowych na chromosomie 14. Dochodzi do nadekspresji protoonkogenu, przekszta│cenia go w onkogen, zaburze± cyklu kom≤rkowego i w efekcie do procesu nowotworowego.
 MATERIAú GENETYCZNY KOM╙RKI Z ZAúO»ENIA NIE JEST NIEZMIENNY
MATERIAú GENETYCZNY KOM╙RKI Z ZAúO»ENIA NIE JEST NIEZMIENNY
TRANSPOZONY - W╩DRUJíCE DNA


⌐ 1997, 1998 Biologia Molekularna w Internecie Webmaster