






OBRÓBKA PRE-mRNA
I tak oto osiągnęliśmy etap pierwotnego transkryptu, który aby stać się matrycą w syntezie białka , musi ulec zasadniczym modyfikacjom. Polegają one na wycięciu intronów oraz ( co jest charakterystyczne dla większości pre-mRNA ) dodaniu na 3' końcu długiego fragmentu tzw. poli A.
1.gif)
Schemat obrazujący procesy
zachodzące na drodze od DNA do mRNA
Poliadenylacja
Znakomita większość produktów reakcji katalizowanej przez polimerazę II RNA zawiera na swoim 3' końcu tzw. ogon poli A, tj. ciąg liczący od 100 do 250 połączonych ze sobą nukleotydów adeninowych. Nie znaczy to jednak, że w większości genów znajduje się obszar bogaty w pary AT. Fragment poli A nie jest efektem transkrypcji a posttranskrypcyjnej modyfikacji, w której udział bierze enzym zwany polimerazą poli A.
Polimeraza poli A dodaje na 3' końcu pierwotnego transkryptu kolejne nukleotydy adeninowe dopiero po zadziałaniu specyficznej nukleazy tj. enzymu tnącego kwas nukleinowy w obrębie jego cząsteczki. Okazuje się bowiem, że ogon poli A nie jest dołączan y do ostatniego nukleotydu wbudowanego na drodze transkrypcji, a do tego, który stał się ostatnim po rozcięciu nici pre-mRNA (zobacz i porównaj: mRNA). Miejsce atakowane przez nukleazę nie jest dowolne, wyznacza je sekwencja: AAUAAA położona w różnej odległości od przeznaczonego m iejsca działania enzymu.
I tak na przykład u ssaków cięcie następuje w odległości 10 - 30 nukleotydów za sekwencją AAUAAA.
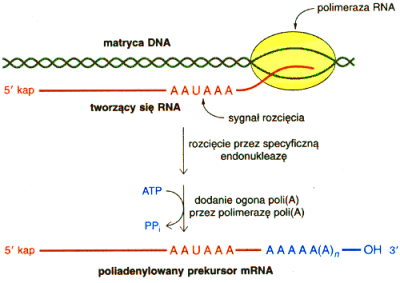
Mechanizm poliadenylacji pre-mRNA (zobacz i porównaj: mRNA)
Istnieją geny zawierające więcej niż jeden sygnał poliadenylacji ( tj.wspomnianą sekwencję ). Oznacza to, że na ich matrycy powstanie kilka pierwotnych transkryptów różniących się długością , a co za tym idzie kilka różnych mRNA. Przykładem alterna tywnej poliadenylacji może być modyfikacja pierwotnego transkryptu szczurzego genu kodującego kalcytoninę.
Gen ten zawiera dwa miejsca poliadenylacji, z których pierwsze preferowane jest w komórkach tarczycy a drugie w mózgu.
Ogon poli A podobnie jak struktura czapeczki chroni cząsteczkę pierwotnego transkryptu przed działaniem nukleaz. Może on mieć również znaczenie w translacji, okazuje się bowiem, że transkrypt pozbawiony ogona poli A jest mniej wydajną matrycą przy sy ntezie białka.
Wycinanie intronów
Zanim powstanie ostateczny, dojrzały mRNA mogący ulec translacji, pre-mRNA musi zostać pozbawiony intronów. Proces wycinania tych niekodujących "wtrętów" zwany jest splicingiem i wykorzystuje jeden wspólny dla eliminacji wszystkich intronów aparat enzymatyczny.
Oznacza to, że introny muszą mieć jakieś wspólne elementy rozpoznawane przez wspomniany aparat splicingowy. Z porównywania sekwencji różnych intronów wynika, że łączą je następujące podobieństwa:
- na 5' końcu zawierają zawsze kolejno: resztę guaninową i uracylową (5'- GU)
- na ich 3' końcu występuje reszta guaninowa poprzedzona resztą adeninową (AG - 3')
- wewnątrz zawierają tzw. miejsce rozgałęzienia, w którym kluczową dla splicingu rolę odgrywa nukleotyd adeninowy.
1.gif)
Sygnały splicingowe
Mechanizm splicingu
Wycinanie intronów, jak każdy proces przeprowadzany przez organizm żywy, jest skomplikowane. Wyróżnić w nim można kilka etapów:
1. Rozcięcie cząsteczki pre-mRNA w miejscu splicingowym 5' (tj. na 5' końcu intronu).W wyniku tej reakcji następuje uwolnienie 5' końca fosforanowego egzonu 1.
2. Atak grupy 2'-OH nukleotydu adeninowego ( z miejsca rozgałęzienia ) na 5' grupę fosforanową nukleotydu guaninowego intronu. Jak wiadomo wiązania między kolejnymi nukleotydami to wiązania fosfodiestrowe tworzone między tzw. 5' grupą fosforanową a gru pą hydroksylową ( -OH ) rybozy. Zwykle do wiązania tego angażowana jest grupa 3'-OH, niemniej jednak inne reszty hydroksylowe m.in. 2' posiadają zdolność do interakcji z resztą fosforanową. I tak nukleotyd adeninowy ( z miejsca rozgałęzienia ) mając grup ę 3'-OH wykorzystaną w "normalnym" wiązaniu fosfodiestrowym do interakcji z resztą fosforanową 5' intronu angażuje grupę 2'-OH. Tworzy się tu wiązanie 2'-5' fosfodiestrowe.
3. Atak uwolnionej grupy 3'-OH egzonu 1 na 5' grupę fosforanową egzonu 2 z uwolnieniem intronu mającego postać lassa.

Mechanizm splicingu pre-mRNA
(kolejność powstawania wiązań)
Spliceosom
Opisane powyżej reakcje nie zachodzą samoistnie. Są one katalizowane przez kompleks przyłączających się kolejno cząstek tworzących spliceosom. W skład aparatu splicingowego wchodzi pięć rodzajów snRNP ( snRNA + białka ): U1, U2, U4, U5 i U6, z który ch każdy pełni odrębną, istotną funkcję:
U1- łączy się z miejscem splicingowym 5'i 3' na zasadzie komplementarności, zawiera on bowiem w swej rybonukleinowej komponencie sekwencję komplementarną do styków egzon-intron;
U2- wiąże miejsce rozgałęzienia;
U6- katalizuje splicing, występuje w kompleksie z U4 i U5.
1.gif)
Sposób działania spliceosomu
Produktem splicingu jest dojrzały mRNA niosący same istotne informacje dotyczące budowy białka ( poza fragmentami skrajnymi, których głównym zadaniem jest stabilizacja transkryptu ). Okazuje się jednak, że na matrycy jednego genu może powstać kil ka różnych mRNA, co poza alternatywną poliadenylacją powodowane jest tzw. alternatywnym splicingiem. Polega on na łączeniu ze sobą różnych egzonów tzn. niekoniecznie wszystkich występujących w pre-mRNA (zobacz i porównaj: mRNA).
Fig.III.2.7. Alternatywny splicing na przykładzie (-tropomiozyny szczura
5'NT i 3'NT - obszary nie ulegające translacji
Nie wiadomo jeszcze dokładnie, dlaczego w różnych tkankach wybierane są odmienne wzory łączenia egzonów. Być może maszyneria obsługująca proces wycinania intronów nie jest identyczna we wszystkich rodzajach komórek tzn. występują pewne subtelne, ale znaczące różnice między spliceosomami poszczególnych tkanek. Możliwe jest także, że podstawowy aparat splicingowy jest uniwersalny, a różne tkanki zawierają dla siebie charakterystyczne czynniki ( białkowe lub RN-owe ) łączące się z pre-mRNA, czego konse kwencją może być ułatwienie bądź utrudnienie działania spliceosomów w różnych miejscach w obrębie pierwotnego transkryptu.
Podsumowując:
1. Pre-mRNA zanim stanie się pełnowartościową matrycą do syntezy białka musi przejść przez proces dojrzewania.
2. Dojrzewanie obejmuje 3 zasadnicze modyfikacje:
- capping ( dodawanie czapeczki )
- poliadenylację podnoszącą stabilność transkryptu;
- splicing, w którym eliminowane są wewnętrzne niekodujące rejony informacyjnego RNA.
3. Alternatywny splicing i poliadenylacja mogą generować różne produkty ( mRNA ) wykrywane w różnych tkankach, co znaczy, że jeden gen może kodować więcej niż jedno białko.
4. Dojrzewanie pre-mRNA jest warunkiem eksportu transkryptu z jądra.
5. Dojrzały mRNA wiąże się z białkami mającymi wpływ na jego stabilność, transport i translację.
 TRANSKRYPCJA
TRANSKRYPCJA
TRANSLACJA


Š 1997, 1998 Biologia Molekularna w Internecie Webmaster