






TRANSKRYPCJA
Transkrypcja to proces, w kt≤rym informacja zawarta w DNA - zapisana w formie sekwencji deoksyrybonukleotyd≤w - przepisana zostaje na jΩzyk rybonukleotyd≤w w pre-mRNA (zobacz i por≤wnaj: mRNA) podczas reakcji katalizowanej przez enzym zwany polimeraz▒ II RNA.
Jak ju┐ wspomniano, ka┐dy z etap≤w ekspresji jest bardzo z│o┐ony. Sama transkrypcja nie jest wyj▒tkiem, dzieli siΩ j▒ bowiem dalej na trzy nastΩpuj▒ce po sobie zdarzenia:
- inicjacjΩ transkrypcji,
- elongacjΩ │a±cucha pre-mRNA (zobacz i por≤wnaj: mRNA),
- terminacjΩ.
Inicjacja transkrypcji
Zanim bΩdzie mo┐na przej╢µ do omawiania tytu│owego zagadnienia, nale┐y zapoznaµ siΩ z budow▒ genu eukariotycznego, kt≤ry jest jednostk▒ koduj▒c▒ bia│ko. Takie ujΩcie znaczenia genu mo┐e byµ bardzo myl▒ce, gdy┐ sugeruje ono, ┐e ca│y gen ( od pierwsze go do ostatniego nukleotydu ) niesie informacjΩ o strukturze bia│ka.
W rzeczywisto╢ci jednak takie wskaz≤wki zawarte s▒ jedynie we fragmentach genu zwanych egzonami. W obrΩbie tej czΩ╢ci genu, kt≤ra zostanie przepisana na mRNA opr≤cz egzon≤w wystΩpuj▒ r≤wnie┐ fragmenty bΩd▒ce najczΩ╢ciej nic nie znacz▒cymi wtrΩtami. Okre╢la siΩ je mianem intron≤w. Jakkolwiek w czasie transkrypcji te "zbΩdne" fragmenty przepisywane s▒ tak samo jak egzony na niµ pierwotnego mRNA ( pre-mRNA ), tak przed zaj╢ciem translacji s▒ one eliminowane.
Gen opr≤cz obszaru transkrybowanego zawiera jeszcze promotor i terminator odpowiednio na swoim 5' i 3' ko±cu, przy czym jedno i drugie nale┐y pojmowaµ raczej jako obszary funkcjonalne a nie jedynie strukturalne.
1.gif)
Schemat budowy genu eukariotycznego
na przyk│adzie genu ssaczego i dro┐d┐owego
( z uwzglΩdnieniem element≤w regulatorowych )
Inicjacja transkrypcji to ten moment, w kt≤rym odbywa siΩ najbardziej precyzyjna kontrola ekspresji. To, czy dany gen w konkretnej kom≤rce zostanie w danym momencie wyra┐ony, tzn. czy zostanie zsyntetyzowane okre╢lone bia│ko, zale┐y w g│≤wnej mierze od tego, czy transkrypcja zostanie zapocz▒tkowana. Oczywi╢cie zdarza siΩ, ┐e transkrypcja zachodzi, ale na dalszych etapach ekspresja zostaje wygaszona, czego skutkiem jest brak syntezy bia│ka. Jednak┐e generalnie inicjacja transkrypcji jest g│≤wnym punk tem kontrolnym ekspresji.
Jako inicjacjΩ okre╢la siΩ zdarzenia zachodz▒ce w obrΩbie promotora, a tak┐e daleko po│o┐onych rejon≤w zwanych enhancerami ( wzmacniaczami ), kt≤re umo┐liwiaj▒ polimerazie II RNA podjΩcie dzia│ania.W przeciwie±stwie do prokariont≤w u polimeraza II RNA wymaga do zapocz▒tkowania reakcji obecno╢ci pewnych bia│ek, kt≤re okre╢la siΩ mianem czynnik≤w transkrypcyjnych tzw. TF-≤w ( ang. transcription factors ). Bia│ka te w okre╢lonym porz▒dku wi▒┐▒ siΩ do DNA w obrΩbie promotora, enhancer≤w b▒d╝ silencer≤w.
To w│a╢nie ich obecno╢µ lub brak w danej kom≤rce ( czy tkance ) decyduje w znacznej mierze o rozpoczΩciu lub te┐ nie transkrypcji konkretnego genu. Opr≤cz bia│ek na transkrypcjΩ mog▒ r≤wnie┐ wp│ywaµ hormony.
Nale┐y tu jednak zaznaczyµ, ┐e jakkolwiek TF-y s▒ istotne w regulacji ekspresji informacji genetycznej, tak najwa┐niejsza jest struktura chromatyny. Wiadomo, ┐e aby zasz│a transkrypcja danego genu musi mieµ do niego dostΩp "aparat enzymatyczny". Je╢ li gen jest │atwo dostΩpny i kom≤rka dysponuje odpowiednimi TF-ami, transkrypcja zachodzi. Je┐eli natomiast gen jest "ukryty" przed polimeraz▒ nic nie pomo┐e obecno╢µ optymalnego zestawu czynnik≤w transkrypcyjnych. To, czy dany gen jest │atwo czy trudno d ostΩpny zale┐y od tego jaka jest struktura chromatyny w rejonie, w kt≤rym wystΩpuje. Im silniejsza jest jej kondensacja tym mniejsza mo┐liwo╢µ przy│▒czenia enzymu. Okazuje siΩ, ┐e w r≤┐nych tkankach r≤┐ne rejony DNA s▒ mocno skondensowane ( sheterochroma tynizowane ). Tak wiΩc, je┐eli wiadomo, ┐e konkretny gen w danej tkance nie ulega transkrypcji z du┐ym prawdopodobie±stwem mo┐na stwierdziµ, ┐e obszar w kt≤rym siΩ lokuje jest niedostΩpny dla aparatu transkrypcyjnego.
Rejony DNA o funkcjach regulacyjnych
Sterowanie transkrypcj▒ - przez zwi▒zanie czynnik≤w bia│kowych czy hormonalnych - mo┐e odbywaµ siΩ z r≤┐nych miejsc na DNA. Miejsca te mog▒ le┐eµ w obrΩbie gen≤w ( promotory ), lub te┐ w odleg│o╢ci kilku tys. nukleotyd≤w ( enhancery, silencery ).
Promotory
Obszar promotorowy genu znajduje siΩ na jego 5' ko±cu i zawiera kilka istotnych rejon≤w rozpoznawanych przez polimerazΩ II RNA ( najbli┐ej miejsca startu transkrypcji ) oraz czynniki transkrypcyjne. Spo╢r≤d wspomnianych rejon≤w najistotniejszym i na jbardziej powszechnym jest kaseta TATA ( tzw. TATA-box ). Jest to 7-nukleotydowa sekwencja po│o┐ona w odleg│o╢ci ok. 25 p.z. od miejsca startu transkrypcji, kt≤ra w pe│ni prezentuje siΩ nastΩpuj▒co: 5'- TATAAAA -3'. Obecno╢µ kasety TATA, choµ niezbΩdna w przypadku prawie wszystkich gen≤w; nie jest wystarczaj▒ca, aby z promotora ruszy│a transkrypcja. Do pe│nej aktywno╢ci promotora niezbΩdne s▒ inne sekwencje wystΩpuj▒ce w rejonie od -110 do -40
( odleg│o╢µ podana w liczbie nukleotyd≤w na lewo od miejsca startu transkrypcji ). Kaseta TATA stanowi tzw. czΩ╢µ rdzeniow▒ promotora. Ponadto promotory gen≤w zawieraj▒ inne - nie tak powszechne - sekwencje, kt≤re odgrywaj▒ znacz▒c▒ rolΩ w poszczeg≤lnych tkankach zaopatrzonych w bia│ka rozpoznaj▒ce owe sekwencje.
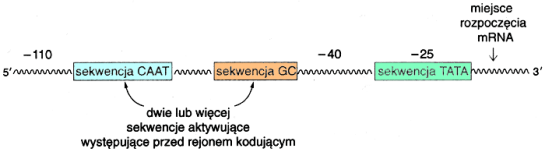
Schemat promotora genu eukariotycznego, uwzglΩdniaj▒cy rejony najbardziej powszechne )
Enhancery i silencery
Enhancery to sekwencje wzmacniaj▒ce aktywno╢µ promotor≤w. Po│o┐one mog▒ byµ nawet w znacznej odleg│o╢ci od genu i zachowuj▒ zdolno╢µ regulacyjn▒ tak┐e po eksperymentalnej zmianie orientacji o 180( wzglΩdem genu. Do enhancer≤w wi▒┐▒ siΩ czynniki trans krypcyjne okre╢lane jako aktywatory, kt≤re odzia│ywuj▒ z polimeraz▒ II RNA i TF-ami promotorowymi. Mo┐liwo╢µ oddzia│ywa± enhancer : promotor mimo odleg│o╢ci pomiΩdzy nimi wynosz▒cej niekiedy kilka tys. p.z. istnieje dziΩki du┐ej elastyczno╢ci nici DNA. Ow a elastyczno╢µ objawia siΩ zdolno╢ci▒ DNA do dowolnego wyginania siΩ.
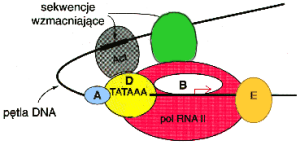
Interakcje enhancer : promotor
Enhancery - rozumiane jako sekwencje -obecne s▒ w ka┐dej kom≤rce ( pamiΩtamy, ┐e DNA we wszystkich kom≤rkach organizmu jest identyczny ), natomiast tylko w niekt≤rych wykazuj▒ aktywno╢µ wzmacniaj▒c▒. Jest to znacz▒cy fakt w regulacji ekspresji info rmacji genetycznej, a bierze siΩ on z r≤┐nic w sk│adzie bia│ek kom≤rkowych poszczeg≤lnych tkanek. A zatem istniej▒ enhancery tkankowo-specyficzne wzmagaj▒ce transkrypcjΩ tylko tam, gdzie obecne s▒ bia│ka wi▒┐▒ce siΩ w ich obrΩbie. Enhancery maj▒ tak┐e is totne znaczenie w regulacji transkrypcji przez hormony steroidowe.
Silencery - ( ang. silence - cisza ), sekwencje s│u┐▒ce wyciszeniu aktywno╢ci promotora; podobnie jak enhancery mog▒ byµ w r≤┐nym stopniu oddalone od genu w obu kierunkach, a tak┐e wystΩpowaµ w jego wnΩtrzu.
Ciekaw▒ sytuacjΩ mo┐na zaobserwowaµ w regulacji transkrypcji gen≤w immunoglobulin ( bia│ek syntetyzowanych jedynie w limfocytach B ). W tym przypadku silencer wbudowany jest w obrΩbie enhancera, a jego znaczenie uwidocznia siΩ w kom≤rkach innych ni┐ limfocyty B. Jest to swoiste zabezpieczenie przed ekspresj▒ gen≤w immunoglobulin zwi▒zane z inaktywacj▒ enhancera.
Zwi▒zki steruj▒ce transkrypcj▒
Regulacja ka┐dego procesu mo┐e odbywaµ siΩ na dwa sposoby: pozytywny b▒d╝ negatywny.
Poniewa┐ w przypadku transkrypcji eukariotycznej lepiej poznano regulacjΩ pozytywn▒ najwiΩcej uwagi zostanie po╢wiΩcone aktywatorom, czyli zwi▒zkom umo┐liwiaj▒cym polimerazie II RNA rozpoczΩcie syntezy nici pre-mRNA (zobacz i por≤wnaj: mRNA).
A. Czynniki transkrypcyjne wi▒┐▒ce siΩ z promotorem.
Eukariotyczna polimeraza II RNA sama w sobie nie jest zdolna do przy│▒czenia siΩ do DNA, a co za tym idzie do rozpoczΩcia transkrypcji. Zanim enzym zostanie zwi▒zany w rejonie startu transkrypcji, do promotora przy│▒czaj▒ siΩ kolejno bia│ka zaliczane do grupy TFII ( II st▒d, ┐e wsp≤│pracuj▒ z polimeraz▒ II ). Kolejno╢µ przy│▒czania siΩ TF-≤w i polimerazy do promotora jest ╢ci╢le okre╢lona.
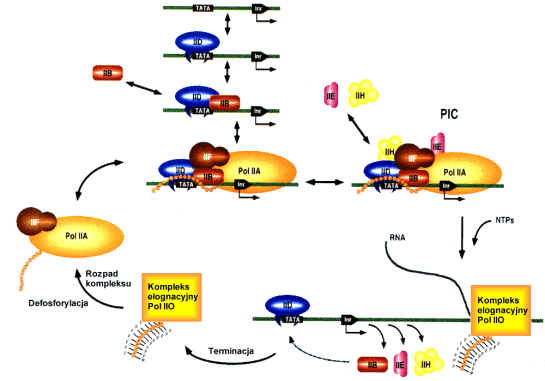
Tworzenie kompleksu preinicjacyjnego
Jako pierwszy do DNA wi▒┐e siΩ TFIID. Jest to kompleks, w kt≤rego sk│ad wchodzi TBP oraz czynniki towarzysz▒ce TBP tzw. TAF ( TBP associated factors ). TBP to bia│ko wchodz▒ce w bezpo╢redni▒ interakcjΩ z DNA w obrΩbie TATA box. Ma ona postaµ siod│a nasadzaj▒cego siΩ na niµ kwasu nukleinowego w taki sposob, ┐e jego kra±cowe fragmenty wciskaj▒ siΩ miΩdzy pary zasad w DNA. Wnikanie pomiΩdzy zasady nici komplementarnych powoduje lekkie ( pierwotne ) rozplecenie helisy, konieczne w procesach takich jak omawiana tu transkrypcja, czy te┐ replikacja.
fig. Model przestrzennej budowy TBP
W dalszej kolejno╢ci przy│▒czeniu do promotora ulega TFIIB i polimeraza II RNA ( w kompleksie z TFIIF ). TFIIF nie kontaktuje siΩ z DNA podobnie jak TFIIE przy│▒czaj▒cy siΩ do powsta│ego kompleksu bia│kowego jako ostatni.
|
Nazwa czynnika |
Funkcje |
|
TFIID - TBP
|
Przy│▒czanie siΩ do TATA box ( rozlu╝nianie helisy DNA ) Umo┐liwianie TFIIB przy│▒czania siΩ do DNA Regulacyjne ( aktywacja, represja ) |
|
TFIIB |
Umo┐liwianie │▒czenia siΩ kompleksu polimerazy i TFIIF z DNA |
|
TFIIF |
Kierowanie polimerazy II RNA do promotora |
|
TFIIE |
Stymulacja aktywno╢ci TFIIH |
|
TFIIH |
Rozplatanie helisy DNA (aktywno╢µ helikazy ) Dostarczanie energii niezbΩdnej do zaj╢cia reakcji ( hydroliza ATP ) Fosforylacja polimerazy ( modyfikacja aktywuj▒ca enzym ) |
Czynniki buduj▒ce wraz z polimeraz▒ II RNA kompleks inicjacyjny
Om≤wiony powy┐ej kompleks nosi nazwΩ podstawowego aparatu transkrypcyjnego i mimo tego, i┐ jest zdolny do przeprowadzenia transkrypcji robi to w znikomym i niewystarczaj▒cym do wyra╝nej ekspresji genu stopniu. Do promotora (opr≤cz wymienionych TF-≤w ) przy│▒czaj▒ siΩ r≤wnie┐ bia│ka takie jak, wspomniane wcze╢niej ssacze Sp1, NF1 oraz szereg innych.
Przyk│adem bia│ek tkankowo- specyficznych s▒ tzw. OTF2A i OTF2B, kt≤re obecne jedynie w limfocytach B - przy│▒czaj▒c siΩ do promotora - umo┐liwiaj▒ ekspresjΩ gen≤w immunoglobulin.B. Czynniki wi▒┐▒ce siΩ z sekwencjami spoza promotora
Dla transkrypcji, kt≤rej efektem by│aby intensywna synteza RNA ( warunek wyra┐enia genu ) niezbΩdna jest obecno╢µ zwi▒zk≤w wzmagaj▒cych transkrypcjΩ podstawow▒. S▒ to aktywatory transkrypcji │▒cz▒ce siΩ z DNA w rejonach enhancerowych. Kierunki oddzi a│ywa± aktywator≤w obrazuje poni┐szy rysunek.

Kierunki oddzia│ywa± aktywator≤w transkrypcji
Jak widaµ, aktywatory mog▒ wp│ywaµ bezpo╢rednio na bia│ka aparatu podstawowego, b▒d╝ te┐ za po╢rednictwem koaktywator≤w. Zwi▒zki aktywuj▒ce mog▒ mieµ r≤┐ny charakter: bia│kowy ( w przewa┐aj▒cej czΩ╢ci ), lub te┐ steroidowy.
Czynniki bia│kowe stale obecne w danej tkance s▒ dla niej charakterystyczne i swoje funkcje w po│▒czeniu z enhancerem mog▒ spe│niaµ tylko w niej.
Czynniki steroidowe, a konkretnie hormony takie jak np.glukokortykoidy pojawiaj▒ siΩ w odpowiedzi na bod╝ce ╢rodowiskowe i s▒ transportowane z miejsc syntezy do kom≤rek docelowych. Hormon, kt≤ry dociera do kom≤rki przeznaczenia wi▒┐e siΩ z receptore m ( nie wszystkie kom≤rki maj▒ receptory na produkowane przez organizm hormony ) i tak powsta│y uk│ad - hormon : receptor - przy│▒cza siΩ do enhancera reguluj▒c w ten spos≤b transkrypcjΩ.
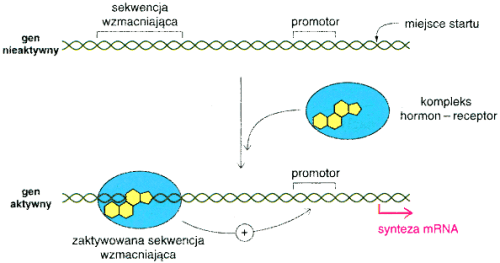
Aktywowanie transkrypcji przez uk│ad hormon : receptor
Podsumowuj▒c:
1. Inicjacja transkrypcji jest momentem najbardziej precyzyjnej kontroli ekspresji genu.
2. Transkrypcja mo┐e zachodziµ na r≤┐nych poziomach:
- podstawowym ( niskim ),
- zaktywowanym ( intensywnym ) w zale┐no╢ci od sk│adu czynnik≤w transkrypcyjnych w danej tkance.
3. Regulacja transkrypcji zale┐na jest do czynnik≤w:
- wewnΩtrznych (struktura chromatyny w rejonie wystΩpowania okre╢lonego genu, obecno╢µ kompletu TF-≤w);
- zewnΩtrznych - odpowied╝ sprowokowana bod╝cami ze ╢rodowiska np. hormonalna.
Synteza nici pre-mRNA (zobacz i por≤wnaj: mRNA)
Po spe│nieniu wszystkich warunk≤w koniecznych do rozpoczΩcia reakcji przez polimerazΩ II RNA, nastΩpuje synteza │a±cucha pre-mRNA ( czyli prekursora mRNA) w kierunku od 5' do 3' ko±ca. Trankrypcja jednego fragmentu katalizowana jest przez jedn▒ cz▒s teczkΩ enzymu.
Budowa polimerazy II RNA
Eukariotyczne polimerazy II RNA sk│adaj▒ siΩ z 8 (12 podjednostek ( tzn. r≤┐nych polipeptyd≤w ), s▒ wiΩc enzymami wysoce z│o┐onymi. Wsp≤ln▒ ich cech▒ jest obecno╢µ podjednostek RPB1, 2, 5, 6, 8, z kt≤rych pierwsze dwie s▒ du┐ymi bia│kami o masach cz▒steczkowych odpowiednio 220 i 140 kDa. Na C-ko±cu ( ko±cu karboksylowym bia│ka ) podjednostki RPB 1 wystΩpuje w zmiennej ( w zale┐no╢ci od organizmu ) liczbie powt≤rze± sekwencja 7-aminokwasowa, kt≤ra okazuje siΩ byµ niezbΩdna do dzia│ania enzymu. Jak do tej pory nie okre╢lono funkcji poszczeg≤lnych podjednostek enzymu eukariotycznego w odr≤┐nieniu od enzymu prokariotycznego, w kt≤rym zdefiniowano rolΩ ka┐dej z podjednostek.
Polimeraza II RNA zlokalizowana jest w nukleoplazmie i opr≤cz syntezy mRNA przeprowadza tam r≤wnie┐ syntezΩ snRNA ( ang. small nuclear RNA ) tj. cz▒steczek, o kt≤rych mowa bΩdzie przy okazji splicingu.

Jak dzia│a polimeraza II RNA?
Jak wiadomo zadaniem polimerazy II RNA jest przepisywanie informacji zawartej w genie ( DNA ) na jΩzyk RNA. Informacja ta ( dotycz▒ca struktury bia│ek ) zapisana jest tylko w jednej nici DNA, w tzw. nici koduj▒cej. Druga niµ - komplementarna - nie niesie informacji o bia│ku, a stanowi matrycΩ dla polimerazy, kt≤ra syntetyzuje pre-mRNA (zobacz i por≤wnaj: mRNA) na zasadzie komplementarno╢ci. Zasada ta polega na wbudowywaniu do powstaj▒cej nici informacyjnego RNA nukleotydu adeninowego (A) je╢li w odczytywanym miejscu na mat rycy obecny jest nukleotyd tyminowy (T), guaninowego (G) je╢li na matrycy jest cytozynowy (C) itd.(Tabela)
|
Nukleotyd na matrycy |
Nukleotyd w│▒czany do pre-mRNA (zobacz i por≤wnaj: mRNA) |
|
A |
U |
|
T |
A |
|
G |
C |
|
C |
G |
A zatem transkrypt jest komplementarny do nici matrycowej i homologiczny z nici▒ koduj▒c▒.
Nale┐y jednak pamiΩtaµ, ┐e jakkolwiek homologami G, C i A w pre-mRNA (zobacz i por≤wnaj: mRNA) s▒ rybonukleotydy nios▒ce te same zasady azotowe, to homologiem T jest rybonukleotyd zawieraj▒cy uracyl (U) a nie tyminΩ.
Por≤wnanie sekwencji pre-mRNA (zobacz i por≤wnaj: mRNA) z sekwencjami nici koduj▒cej i matrycowej genu.
( kolorem czerwonym oznaczono niµ koduj▒c▒, a zielonym niµ matrycow▒ )
5'- A A T C G G C A T G C C A T G G C C T T G C G C T A - 3' Gen
3'- T T A G C C G T A C G G T A C C G G A A C G C G A T - 5'
5'- A A U C G G C A U G C C A U G G C C U U G C G C U A - 3' pre-mRNA
Zgodnie z tym co zosta│o powiedziane, enzym syntetyzuj▒cy pre-mRNA sun▒c po DNA dobiera odpowiednie rybonukleotydy i przy│▒cza je kolejno w kierunku 5'- 3'( tzn. do wolnej grupy 3'OH pierwszego nukleotydu przy│▒czona zostaje 5' grupa fosforanowa kol ejnego nukleotydu ). Kierunek ten obowi▒zuje w ka┐dej reakcji polimeryzacji kwas≤w nukleinowych i wynika z mechanizmu tworzenia wi▒za± miΩdzy s▒siaduj▒cymi ze sob▒ nukleotydami - wi▒za± fosfodiestrowych. ( Patrz. Chemiczne podstawy biologii molekularnej. ) Polimeraza RNA jest enzymem wprowadzaj▒cym wiΩcej pomy│ek od polimerazy DNA poniewa┐ nie wykazuje w│a╢ciwo╢ci korekcyjnych, st▒d przepisanie informacji z DNA na RNA nie jest tak wierne jak z DNA na DNA w procesie replikacji. Brak system≤w korekcyjnych w transkrypcji nie niesie ze sob▒ dla kom≤rki powa┐nych konsekwencji, gdy┐ nieprawid│owa cz▒steczka mRNA jest jedn▒ w╢r≤d wielu prawid│owych.
Synteza │a±cucha pre-mRNA (zobacz i por≤wnaj: mRNA) zaczyna siΩ zwykle od zwi▒zania przez enzym nukleotydu guaninowego lub adeninowego, do kt≤rych przy│▒czane s▒ kolejne ,, cegie│ki''.
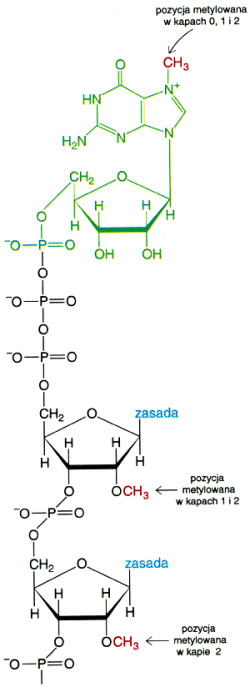 Struktura kap≤w |
Bardzo charakterystycznym dla eukariont≤w zjawiskiem jest tworzenie w pierwotnym transkrypcie struktury czapeczki ( ang. cap ) wp│ywaj▒cej na podniesienie stabilno╢ci pre-mRNA (zobacz i por≤wnaj: mRNA). Proces ten zachodzi r≤wnolegle z transkrypcj▒ i jest jednym z etap ≤w obr≤bki pre-mRNA (zobacz i por≤wnaj: mRNA)( inne rodzaje obr≤bki zachodz▒ po zsyntetyzowaniu ca│ej nici pierwotnego mRNA i zostan▒ om≤wione osobno ). Transkrypt uposa┐ony w czapeczkΩ jest mniej wra┐liwy na dzia│anie nukleaz i fosfataz tj. enzym≤w odpowiedzialnych za degradacjΩ kwasu nukleinowego. Rozpoznanie cap jest tak┐e istotne w inicjacji translacji. Mechanizm tworzenia czapeczki obejmuje 3 zasadnicze etapy: - modyfikacjΩ trifosforanu 5' ko±ca ( patrz. "Struktura bia│ek i kwas≤w nukleinowych" ) przez uwolnienie jednej z reszt fosforanowych na drodze hydrolizy; - przy│▒czenie GTP wi▒zaniem 5'-5'- trifosforanowym; - metylacja guaniny w pozycji N7 tj. przy│▒czenie reszty metylowej (-CH3 ) do atomu azotu 7-go w cz▒steczce guaniny w wyniku reakcji:
reakcja metylacja reszt cukrowcowych (rybozy) kolejnych nukleotyd≤w mo┐e wygenerowaµ kolejne czapeczki (1, 2). |
Wyd│u┐anie │a±cucha pre-mRNA (zobacz i por≤wnaj: mRNA) ko±czy siΩ w miejscach terminacji, kt≤re dosyµ dok│adnie poznane s▒ u Procaryota, natomiast nadal s│abo scharakteryzowane u organizm≤w wy┐szych.
Podsumowanie:
1. Transkrypcj▒ rz▒dzi zasada komplementarno╢ci.
2. Wyd│u┐anie nici pre-mRNA (zobacz i por≤wnaj: mRNA) zachodzi w kierunku 5'-> 3'.
3. R≤wnolegle z elongacj▒ pierwotnego transkryptu zachodzi modyfikacja jego 5' ko±ca tj. tworzenie struktury czapeczki.
4. Produktem reakcji katalizowanej przez polimerazΩ II RNA jest pre-mRNA tzn. cz▒steczka zawieraj▒ca introny.
 EKSPRESJA INFORMACJI GENETYCZNEJ
EKSPRESJA INFORMACJI GENETYCZNEJ
OBR╙BKA PRE-mRNA


⌐ 1997, 1998 Biologia Molekularna w Internecie Webmaster