|
|
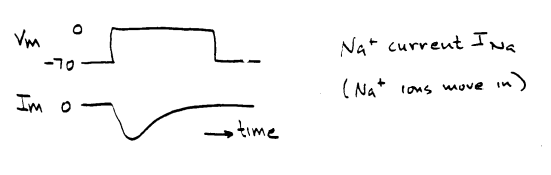
HODGKIN AND HUXLEY'S EXPERIMENTSMany of the details of the basis of nerve electrical activity came from a series of experiments in the early 1950's, which led to the awarding of the Nobel prize to the english physiologists A. L. Hodgkin and A. F. Huxley. As you know, it is now possible to record directly the currents through individual channels, and the approaches used to analyze individual channel behavior are different from Hodgkin and Huxley's analysis of conductance properties of many channels acting in concert. Nevertheless, Hodgkin and Huxley's analysis is a reasonable way of introducing the macroscopic behavior of nerve membrane conductances. If voltage is the governing factor, one needs a method that allows control of the membrane voltage, so that it is possible to explore the voltage dependence of membrane properties. The "voltage clamp" is a way of doing this. Without going into the details, one controls the membrane voltage electronically, and measures the currents across the membrane produced by ion movements. For example, suppose the membrane voltage is forced to change in a stepwise fashion from -70 mV to 0 mV:
The current measured is produced by the movement of ions, but it does not tell one directly what types of ions moved. An outward current could be produced by potassium or sodium moving out of the cell, or by chloride moving into the cell. To determine which ions are responsible, one can either change the concentrations of ions surrounding the membrane, or apply chemicals that interfere with transmembrane movements of specific types of ions. If one takes away the sodium bathing the axon, and produces the same depolarization as above, the current is altered:
Now put the sodium back in, and block the movement of potassium by a drug:
By such experiments, Hodgkin and Huxley determined that, when a nerve membrane is depolarized above a threshold value, there is a transient inward or negative current produced by inward movement of sodium, followed by a more slowly developing and sustained current in the opposite (outward or positive) direction, produced by efflux of potassium. The influx of positive sodium ions depolarizes the cell. This influx is transient. It is followed by an increase in the efflux of positive potassium ions, and this repolarizes the cell. It is not really essential that the potassium conductance increase when the cell is depolarized, because the potassium conductance is already high (the resting cell has a high potassium conductance). A cell in which the potassium conductance remained constant at the resting value would repolarize once the transient increase of sodium conductance was over. However, the increased potassium conductance that takes place upon depolarization hastens repolarization. QUANTITATIVE TREATMENT OF THE CONDUCTANCE CHANGESHodgkin and Huxley applied Ohm's Law to ionic current: I = g V The sodium current, which they called INa, will depend on the membrane's conductance for sodium, gNa, and on the difference between the existing membrane potential Vm, and the membrane potential at which there would be no net electrochemical driving force on sodium. The membrane potential at which there would be no electrochemical driving force on sodium is the Nernst equilibrium potential, ENa. Thus,
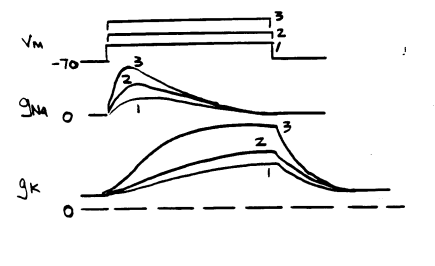
They wrote a similar equation for potassium current: IK = gK(Vm - EK) They observed that Vm is constant when the voltage is controlled by the voltage clamp apparatus, and ENa and EK should not change. So the fact that the currents were functions of time indicated that conductances to sodium and potassium were functions of time, as well as of voltage. To show this, they depolarized axons to a variety of potentials and measured the sodium and potassium currents as functions of time. They divided each sodium current by the term (Vm - ENa) and each potassium current by the term (Vm - EK). This gave them the conductance for each ion as a function of time at each voltage: Depolarization produces a transient increase in gNa, and a slower and sustained increase in gK. Immediately after a step depolarization, gNa>>gK, but after the cell has been depolarized for a time, gK>>gNa. Both gNa and gK are functions of voltage and time: gNa = f(V,t) gK = f'(V,t) Hodgkin and Huxley introduced the variables m, h, and n to help them express the time and voltage dependence of the two conductances. I will just consider the way they described the sodium conductance, because it is the most difficult. They defined two arbitrary variables, m and h. Each of these is a function of voltage and time. They used the equation: gNa = m3h*GNa In this equation, GNa is the maximum sodium conductance of the axon membrane. Sodium traverses the membrane through protein channels in the membrane, and these can open and close. GNa is the conductance for sodium that is seen if all sodium channels are open. The variables m and h each change between 0 and 1 as functions of time and voltage. The product m3h represents the fraction of the total sodium conductance at any given time. The following graph illustrates the values reached by m and h when the membrane is held at various potentials for long times:
Next, they described the time dependence of m and h. If the cell is kept at one voltage for a long time, then voltage is stepped quickly to a new voltage, m and h change to new steady state values, where the steady state values are given by the above graph.
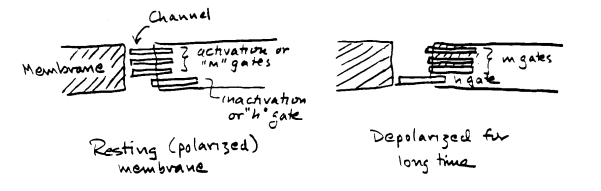
At this point, it is not surprising if the concept of arbitrary variables is disturbing to you. Hodgkin and Huxley suggested a simple physical model which might or might not represent reality, but which helps one see how the time and voltage dependence of currents might arise. The sodium channel behaves as if it has two sets of gates. One set, the "activation" gates, open rapidly when the cell is depolarized above a threshold voltage. The other gates, the "inactivation" gates, close slowly when the cell is depolarized. Suppose each sodium channel has 3 activation gates, called "m" gates, and 1 inactivation gate, called an "h" gate.
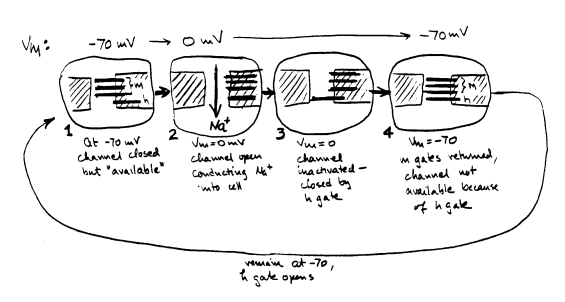
For a single channel, the variable m described by Hodgkin and Huxley is the probability that 1 m gate will be in the open position. m cubed is the probability that all three m gates will be open. When m is near 1, as it is when voltage is near 0 mV, there is a very high probability that all 3 m gates are open. When m equals 0, as it is when voltage is at the resting potential, there is a high probability that the 3 m gates are all shut. For a single channel, the variable h is the probability that the single h gate will be in the open position. When h equals 1 (near the resting potential), the h gate is likely to be open. When h equals 0, as it does in the steady state at depolarized voltages, the h gate is likely to be shut. The product m3h is the probability that all of the gates are open, so that the channel can conduct sodium across the membrane. For a large population of channels, the conductance gNa is equal to the conductance seen when all channels are open (GNa) times the probability that any one channel is open: gNa = m3h GNa Below is an attempt to define the different possible states of these gates at different times after a sudden depolarization to a voltage above threshold, leading to an action potential:
|
|
Copyright 1998, Joe Patlak and Ray Gibbons, Department of
Physiology, University of Vermont. |
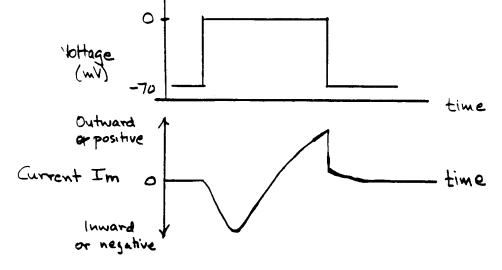 The
terms inward or negative for current in one direction and outward or positive
for current in the opposite direction are conventions. An inward movement of positive ions
is defined as negative or inward current, whereas outward movement of positive ions
produces positive or outward current. Electrically, inward movement of negative ions
cannot be distinguished from outward movement of positive ions, so inward movement of
negative ions produces a positive or outward current.
The
terms inward or negative for current in one direction and outward or positive
for current in the opposite direction are conventions. An inward movement of positive ions
is defined as negative or inward current, whereas outward movement of positive ions
produces positive or outward current. Electrically, inward movement of negative ions
cannot be distinguished from outward movement of positive ions, so inward movement of
negative ions produces a positive or outward current. 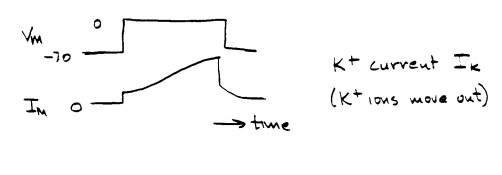
 INa
= gNa(Vm - ENa)
INa
= gNa(Vm - ENa) 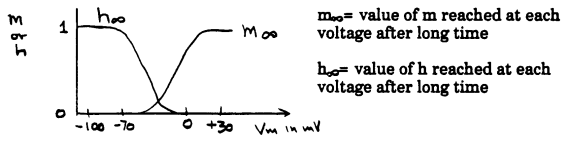
 An
all important fact, however, is that m changes quickly and h changes more slowly. For
example, the following represents the approximate time courses of change of m and h if the
voltage is stepped from -70 mV to 0 mV.
An
all important fact, however, is that m changes quickly and h changes more slowly. For
example, the following represents the approximate time courses of change of m and h if the
voltage is stepped from -70 mV to 0 mV.