






REKOMBINOWANE DNA W PRZEMYªLE.
Bardziej prozaicznym zastosowaniem in┐ynierii bia│ek mo┐e byµ ulepszanie enzym≤w stosowanych przy produkcji detergent≤w np. subtylizyny. Nowy jej wariant to tzw. cz▒steczka drugiej generacji , kt≤ra nie jest dezaktywowana przez wybielacze. PodjΩte s▒ tak┐e prace nad cz▒steczk▒ trzeciej generacji , kt≤ra wykazywa│aby zmniejszon▒ wra┐liwo╢µ na temperaturΩ.
Ponadto nowe technologie znajduj▒ zastosowanie w :
- syntezie biopolimer≤w
- produkcji opakowa±.
Thaumatyna - ro╢linne bia│ko tysi▒c razy s│odsze ni┐ sacharoza mo┐e mieµ niesamowite znaczenie jako potencjalny dodatek do produkt≤w spo┐ywczych , zastΩpuj▒c chemiczne s│odziki takie jak np. sacharyna. Gen koduj▒cy thaumatynΩ zosta│ sklonowany w wektorze dro┐d┐owym pod silnym promotorem. Bia│ko jest produkowane w bardzo du┐ej ilo╢ci , co w warunkach naturalnych nie jest osi▒galne.
Renina - bia│ko u┐ywane w wielkich ilo╢ciach w przemy╢le serowarskim. Do genu bia│ka zosta│a dodana sekwencja sygna│owa , kt≤ra powoduje , ┐e jest ono wydzielane bardzo intensywnie we w│a╢ciwej formie przez dro┐d┐e.
Udoskonalono pewne cechy ro╢lin korzystne przemys│owo np. stworzono odmiany wysokoskrobiowe czy odmiany o po┐▒danym sk│adzie skrobii - zmiana proporcji amylazy i amylopektyny nadaj▒ca przezroczysto╢µ , zmiana procentu kwas≤w t│uszczowych nienasyconych.
TRANSGENICZNE ROªLINY I ZWIERZ╩TA - STRACH MA WIELKIE OCZY.
Organizm transgeniczny - organizm wy┐szy , do genomu kt≤rego wprowadzono nowy gen , heterologiczny (z innego organizmu) w taki spos≤b , aby znalaz│ siΩ on zar≤wno w kom≤rkach somatycznych jak i kom≤rkach pasma p│ciowego dziΩki czemu podlega dziedziczeniu. Jest to trwa│a modyfikacja genetyczna.
Technika uzyskiwania takich organizm≤w r≤┐ni siΩ w zale┐no╢ci od tego czy mamy do czynienia z ro╢lin▒ czy zwierzΩciem.
ROªLINY.
Ich wielka plastyczno╢µ rozwojowa i genetyczna jest spowodowana faktem , ┐e kom≤rki tkanek tw≤rczych , odpowiadaj▒ce kom≤rkom embrionalnym , wystΩpuj▒ w ro╢linie przez ca│e ┐ycie a i niekt≤re kom≤rki zr≤┐nicowane do╢µ │atwo ulegaj▒ odr≤┐nicowaniu - s▒ totipotentne. Ponadto │atwo ulegaj▒ transformacji. Nie jest to jednak r≤wnoznaczne ze stabiln▒ ekspresj▒. NajczΩ╢ciej efekt transgenu ga╢nie po paru pokoleniach poniewa┐ istniej▒ aktywne mechnizmy eliminuj▒ce efekt zak│≤cenia r≤wnowagi. Inn▒ niedogodno╢ci▒ jest brak mo┐liwo╢ci u ro╢lin celowania w okre╢lone miejsce w genomie poniewa┐ homologiczna rekombinacja jest na bardzo niskim poziomie.
Metody transgenizacji :
- Naturalny system wprowadzania DNA - plazmidy Agrobacterium jako wektory DNA dla ro╢lin dwuli╢ciennych.
Plazmidy Ti z Agrobacterium tumefaciens koduj▒ geny odpowiedzialne za syntezΩ pochodnych aminokwas≤w tzw. opin , kt≤re bakterie wykorzystuj▒ jako ╝r≤d│o wΩgla. Poza tym na plazmidzie s▒ geny steruj▒ce katabolizmem opin , warunkuj▒ce transfer plazmidu i jego replikacjΩ. CzΩ╢µ tego plazmidu tzw. T-DNA d│ugo╢ci oko│o 23 kb po zaka┐eniu ulega integracji z genomem ro╢liny i w│a╢nie zamiast tego odcinka mo┐emy wstawiµ obce DNA. Na obu jego ko±cach le┐▒ kr≤tkie powt≤rzenia (24-25 bp) niezbΩdne do integracji z genomem. W procesie przenoszenia T-DNA do kom≤rki ro╢linnej uczestnicz▒ tak┐e geny plazmidowego regionu vir i pewne geny zlokalizowane na chromosomie bakterii.
- Mo┐liwa jest tak┐e bezwektorowa transfekcja kom≤rek ro╢linnych.
Inkubacja ro╢linnych protoplast≤w ze zlinearyzowanym plazmidem DNA zawieraj▒cym marker selekcyjny w warunkach umo┐liwiaj▒cych pobranie DNA np. z glikolem polietylenowym w alkalicznym ╢rodowisku czy elektroporacja (kr≤tkotrwa│e , wysokonapiΩciowe impulsy elektryczne). Inny spos≤b to mikroiniekcja - bezpo╢rednie wstrzykniΩcie DNA a tak┐e
zastosowanie ôarmatki genowejö - wstrzeliwanie drobnych kuleczek wolframowych op│aszczonych DNA. U┐ywa siΩ r≤wnie┐ liposomy (lipidowe pΩcherzyki zapewniaj▒ce ochronΩ przed nukleazami) ze zrekombinowanym DNA , kt≤re ulegaj▒ fuzji z protoplastami.
Sukcesy:
- ZwiΩkszanie trwa│o╢ci ro╢lin dziΩki ingerencji w ich metabolizm np. stworzenie niegnij▒cego pomidora poprzez wykorzystanie strategii antysensu tzn. transformacja antysensownym RNA . Gen antysensownego RNA ma sekwencjΩ komplementarn▒ do genu , kt≤rego ekspresjΩ zamierzamy wy│▒czyµ - w tym przypadku do genu poligalakturonazy. Oba geny ulegaj▒ transkrypcji a wiΩc powstaje mRNA i antysensowny mRNA. ú▒cz▒ siΩ one tworz▒c dupleksy uniemo┐liwiaj▒c syntezΩ bia│ka. Mo┐liwa jest tak┐e ingerencja w system syntezy etylenu przy pomocy strategii kosupresji. Wprowadza siΩ do genomu gen natywny dodatkowy , zamiast wzmo┐onej ekspresji obserwuje siΩ efekt odwrotny a┐ do wygaszenia. Uzyskano w ten spos≤b niewiΩdn▒ce go╝dziki.

- Wykorzystanie ro╢lin jako bioreaktor≤w do produkcji bia│ek heterologicznych spowodowane jest szeregiem zalet , m.in. tanimi kosztami i du┐▒ ilo╢ci▒ produkowanego bia│ka. W ten spos≤b produkowane s▒ interleukiny , cytokiny , hormony wzrostu , przeciwcia│a.
- Uzyskanie nowych odmian - nowych kolor≤w , kszta│t≤w , w│a╢ciwo╢ci wzrostu np. ro╢lin ozdobnych m.in. pomara±czowe petunie (transformacja z udzia│em Agrobacterium genem ze szlaku produkcji antocyjaniny z kukurydzy) czy przekszta│cenie ziemniaka w ro╢linΩ ozdobn▒.


- Uodpornienie ro╢lin na infekcje co daje zwiΩkszone plony , lepszy wzrost i wy┐sz▒ jako╢µ poprzez zainfekowanie ich szczepem wirusowym powoduj▒cym │agodniejszy efekt chroni przed infekcj▒ przez bardziej zjadliwy szczep lub transgenizacja genami koduj▒cymi bia│ko p│aszcza danego wirusa np. wirusa mozaiki tytoniowej.
- Ochrona przed owadami np. dziΩki ekspresji toksyny bakteryjnej Bacillus thuringiensis chroniona jest bawe│na lub dziΩki transgenicznej ekspresji inhibitora proteaz serynowych - tyto±.
- Odporno╢µ ro╢lin uprawnych na herbicydy uzyskiwana na trzy sposoby : nadprodukcja enzymu inaktywowanego przez dany herbicyd , ekspresja zmutowanego enzymu , na kt≤ry zwi▒zek nie dzia│a albo ekspresja enzymu , kt≤ry detoksyfikuje herbicyd.
ZWIERZ╩TA.
Transgeniczne zwierzΩta to organizmy , kt≤re maj▒ obcy DNA wbudowane trwale do kom≤rek rozrodczych - kom≤rek linii p│ciowej.
Uzyskuje siΩ je trzema sposobami :
- dziΩki mikroiniekcji in vitro DNA do jednego z przedj▒drzy zap│odnionej kom≤rki jajowej przed pierwszym jej podzia│em ,
1.gif)
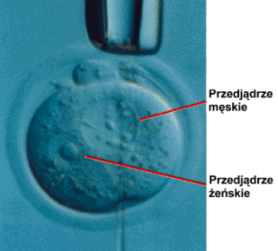
- przez infekcjΩ wczesnego zarodka zrekombinowanym wektorem pochodzenia wirusowego ,
- poprzez modyfikacjΩ genetyczn▒ pierwotnych kom≤rek wΩz│a zarodkowego i wprowadzenie ich do zarodka w stadium blastocysty, poniewa┐ kom≤rki wΩz│a zarodkowego s▒ zdolne do r≤┐nicowania siΩ we wszystkie typy kom≤rek,
1.gif)
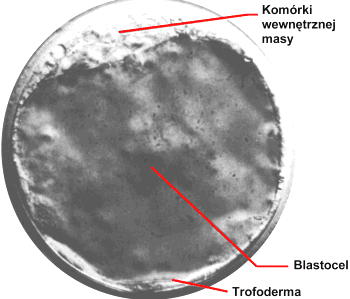
doros│a mysz poddana wcze╢niej takiemu zabiegowi bΩdzie chimer▒ - bΩdzie z│o┐ona z kom≤rek wΩz│a zarodkowego oryginalnego zarodka i z kom≤rek transgenicznych , dalsze krzy┐≤wki pozwalaj▒ uzyskaµ myszy homozygotyczne pod wzglΩdem transgenu.
1.gif)
Pierwszym zwierzΩciem transgenicznym by│a mysz z genem hormonu wzrostu szczura.
Pewien procent transgen≤w w│▒cza siΩ we w│a╢ciwej pozycji genomowej w wyniku homologicznej rekombinacji. Pozwala to na celowe uszkodzenie badanego genu i uzyskanie tzw. myszy transgenicznych z`knock-out`owanych dziΩki kt≤rym mo┐liwe staje siΩ badanie roli pewnych gen≤w. 1.gif)
1.gif)
Myszy takie mog▒ byµ u┐ywane jako modele zwierzΩce ludzkich chor≤b , co ma nieocenion▒ warto╢µ w poznawaniu procesu przebiegu a tak┐e projektowaniu metod leczenia. Transgeniczne myszy nie posiadaj▒ce funkcjonalnego genu rag-2 (aktywatora rekombinazy 2) nie produkuj▒ limfocyt≤w T i B i s▒ odpowiednikiem SCID. Uda│o siΩ tak┐e stworzyµ myszy defektywne w wytwarzaniu interferonu , interleukin i in.
Przeprowadzono podobne do╢wiadczenia na kr≤likach , ╢winiach i owcach. Istotnym ograniczeniem by│a czΩsto╢µ z jak▒ powstawa│y transgeniczne zwierzΩta poniewa┐ tylko 1 na 200 pr≤b mikroiniekcji do jaja dawa│a takie zwierzΩ. To jest jakie╢ 10 do 15 razy rzadziej ni┐ w przypadku myszy.
Jedn▒ z dziedzin biotechnologii , obecnie bardzo intensywnie rozwijan▒ jest klonowanie zarodk≤w ssak≤w. DziΩki temu mo┐liwe jest uzyskanie wielu identycznych osobnik≤w. Wzbudza to ogromne zainteresowanie m.in. hodowc≤w maj▒cych nadzieje na stworzenie jak najwiΩkszej liczby zwierz▒t o po┐▒danych cechach.
Przeprowadza siΩ to wieloma metodami :
- Metod▒ izolacji blastomer≤w.
W miarΩ kolejnych podzia│≤w zygoty powstaje wiele kom≤rek potomnych - blastomer≤w. Pocz▒tkowo ka┐dy z nich jest taki sam i teoretycznie posiada mo┐liwo╢µ utworzenia wszystkich pozosta│ych kom≤rek. Znaczy to , ┐e w tym stadium z ka┐dego blastomeru mo┐e powstaµ ca│y organizm. Do╢wiadczalnie zosta│o to potwierdzone dla stadium nie przekraczaj▒cego o╢miu blastomer≤w. Jest to tak┐e uzale┐nione od gatunku.
- Metod▒ agregacji blastomer≤w.
Jest to w pewnym sensie odmiana metody poprzedniej. Otaczaj▒c pojedynczy blastomer innymi zwiΩksza siΩ szansΩ wytworzenia wΩz│a zarodkowego w og≤le (niezbΩdna jest obecno╢µ pewnej masy kom≤rkowej) a zarazem utworzenia go z centralnie ulokowanego blastomeru. Nie znalaz│a praktycznego zastosowania.
- Metod▒ bisekcji zarodk≤w.
Najszerzej stosowana. Polega na mikrochirurgicznym dzieleniu zarodk≤w (wczesnych zarodk≤w , morul , blastocyst) na po│owΩ.
- Metod▒ transplantacji j▒der kom≤rek zarodkowych.
Jest to jedyna metoda pozwalaj▒ca na uzyskanie klon≤w licz▒cych wiΩksz▒ liczbΩ osobnik≤w. Usuwane s▒ oba przedj▒drza zygoty lub chromosomy z nie zap│odnionych oocyt≤w a nastΩpnie wprowadzane s▒ na ich miejsce j▒dra z blastomer≤w zarodk≤w. Wprowadzenie uzyskuje siΩ r≤┐nymi metodami : mikrochirurgiczn▒ transplantacj▒ , metod▒ fuzji kom≤rkowych czego odmian▒ jest elektrofuzja itd.
Wszystkimi wy┐ej wymienionymi metodami podejmowano pr≤by klonowania przedstawicieli takich gatunk≤w jak myszy , kr≤liki , owce i in. Nale┐y jednak podkre╢liµ , ┐e by│y to manipulacje kom≤rkami niezr≤┐nicowanymi - kom≤rkami embrionalnymi a wiΩc zachowuj▒cymi pewn▒ naturaln▒ totipotentno╢µ.
Zupe│nie odrΩbn▒ metod▒ s▒ pr≤by klonowania z wykorzystaniem doros│ych , zr≤┐nicowanych kom≤rek ssaczych. Najs│awniejsza owca ╢wiata - Dolly powsta│a w ten w│a╢nie spos≤b. Po raz pierwszy uda│o siΩ sklonowaµ organizm korzystaj▒c z kom≤rki doros│ego ssaka. Materia│ genetyczny doros│ej , somatycznej kom≤rki jednej owcy wprowadzono do cytoplazmy kom≤rki jajowej pozbawionej j▒dra drugiej owcy i implantowano do macicy trzeciej. W tym przypadku nale┐y jednak zachowaµ nieco sceptycyzmu. Przede wszystkim dlatego , ┐e eksperymentu tego nie uda│o siΩ jeszcze nikomu powt≤rzyµ. Ponadto pr≤ba ta uda│a siΩ tylko raz na niemal trzysta. Poniewa┐ wiadomo sk▒din▒d , ┐e z podobn▒ czΩsto╢ci▒ wystΩpuj▒ w tkance , z kt≤rej pobrano kom≤rki , kom≤rki niezr≤┐nicowane a tak┐e , ┐e owca , od kt≤rej je pobrano by│a w ci▒┐y , sklonowane zwierzΩ mo┐e byµ pomy│k▒ lub przypadkiem. Technika jak▒ zastosowano wi▒┐e siΩ ponadto ze sporym ryzykiem deformacji rozwojowych a tak┐e jej istotn▒ wad▒ jest bardzo niska wydajno╢µ. Daleka wiΩc jeszcze droga do sukcesu w tej dziedzinie. Jednak wy╢cig ju┐ siΩ zacz▒│.
 REKOMBINOWANE DNA W MEDYCYNIE I FARMACJI
REKOMBINOWANE DNA W MEDYCYNIE I FARMACJI
PROJEKTY - KROK W XXI WIEK ?


⌐ 1997, 1998 Biologia Molekularna w Internecie Webmaster